The Chemical Senses: Gustation and Olfaction
我们的化学感官——Gustation (味觉) 和 Olfaction (嗅觉)——是进化上最古老的感觉系统。它们的最基本任务是探测环境中的化学物质,这对于生存至关重要,能够帮助生物区分营养源(如糖类)与潜在毒素(如许多生物碱),并引导寻找配偶等社会行为。虽然在较低处理水平上味觉与嗅觉是相互独立的系统,但神经系统通过整合这两者的信息,才能产生我们日常体验中的“风味 (Flavor)”。这种信息的融合通常发生在皮层的高级处理阶段。
Gustation (味觉)
The Organs and Structure of Taste
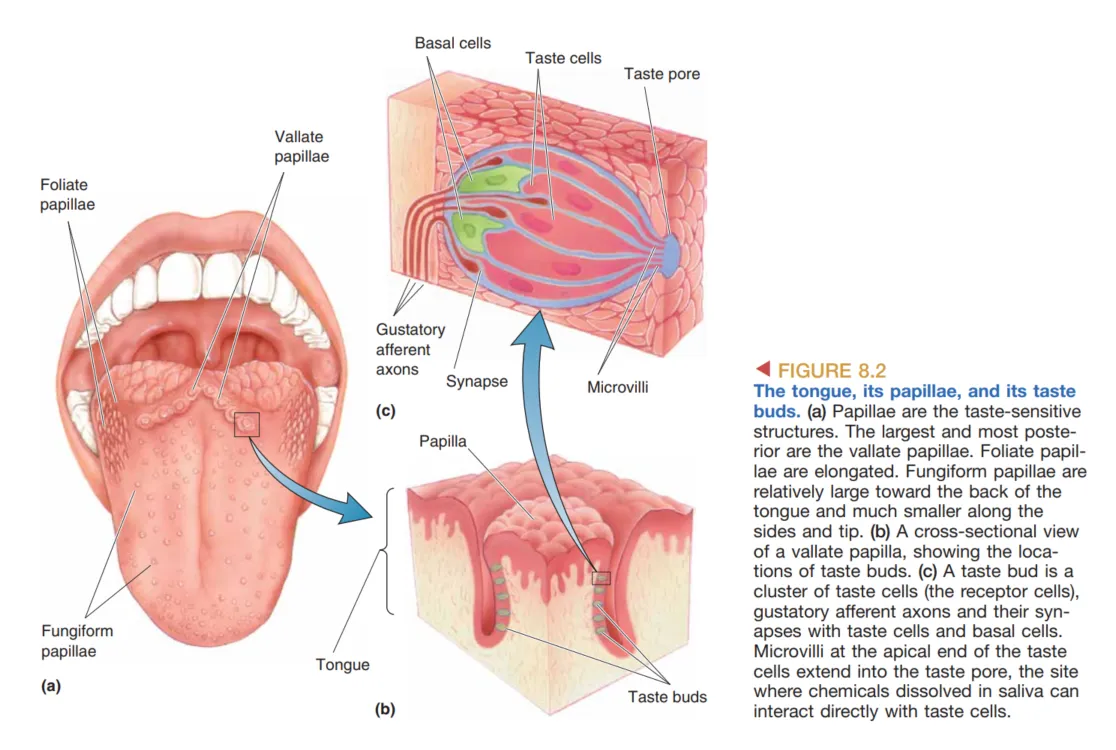
主要的味觉器官是 Tongue (舌),但味觉敏感性并不局限于此。咽 (Pharynx)、腭 (Palate) 和会厌 (Epiglottis) 同样含有味觉感受器。舌表面布满了名为 Papillae (乳突) 的小凸起,根据形状可分为长圆锥形、叶状或蕈状。每个乳突包含一个或多个 Taste Buds (味蕾),而每个味蕾则由 50 至 150 个 Taste Receptor Cells (味觉受体细胞) 像橙子瓣一样排列组成。
值得注意的是,味觉受体细胞在组织学标准下不是神经元,但它们与味蕾底部的 Gustatory Afferent Axons (味觉传入纤维) 形成化学突触。这些细胞的寿命非常短暂(约 2 周),会经历持续的生长、死亡与再生。其顶端具有名为 Microvilli (微绒毛) 的细长延伸,通过 Taste Pore (味孔) 伸出,与舌表面接触的化学物质相互作用。
The Basic Tastes and Survival Significance
目前公认有 5 种基本味觉。Saltiness (咸) 主要是为了平衡电解质;Sourness (酸) 指示 pH 值及可能的腐败;Sweetness (甜) 是碳水化合物等能量物质的指标;Umami (鲜) 指示蛋白质的摄入;而 Bitterness (苦) 则作为一种防御机制,警示环境中可能存在的毒素。味觉感知的复杂性来自于多种基本味觉的组合、口感(Texture)以及嗅觉的协同作用。
Mechanisms of Taste Transduction
不同的味觉物质(Tastants)通过差异化的转导机制产生 Receptor Potential(受体电位),进而触发递质释放。
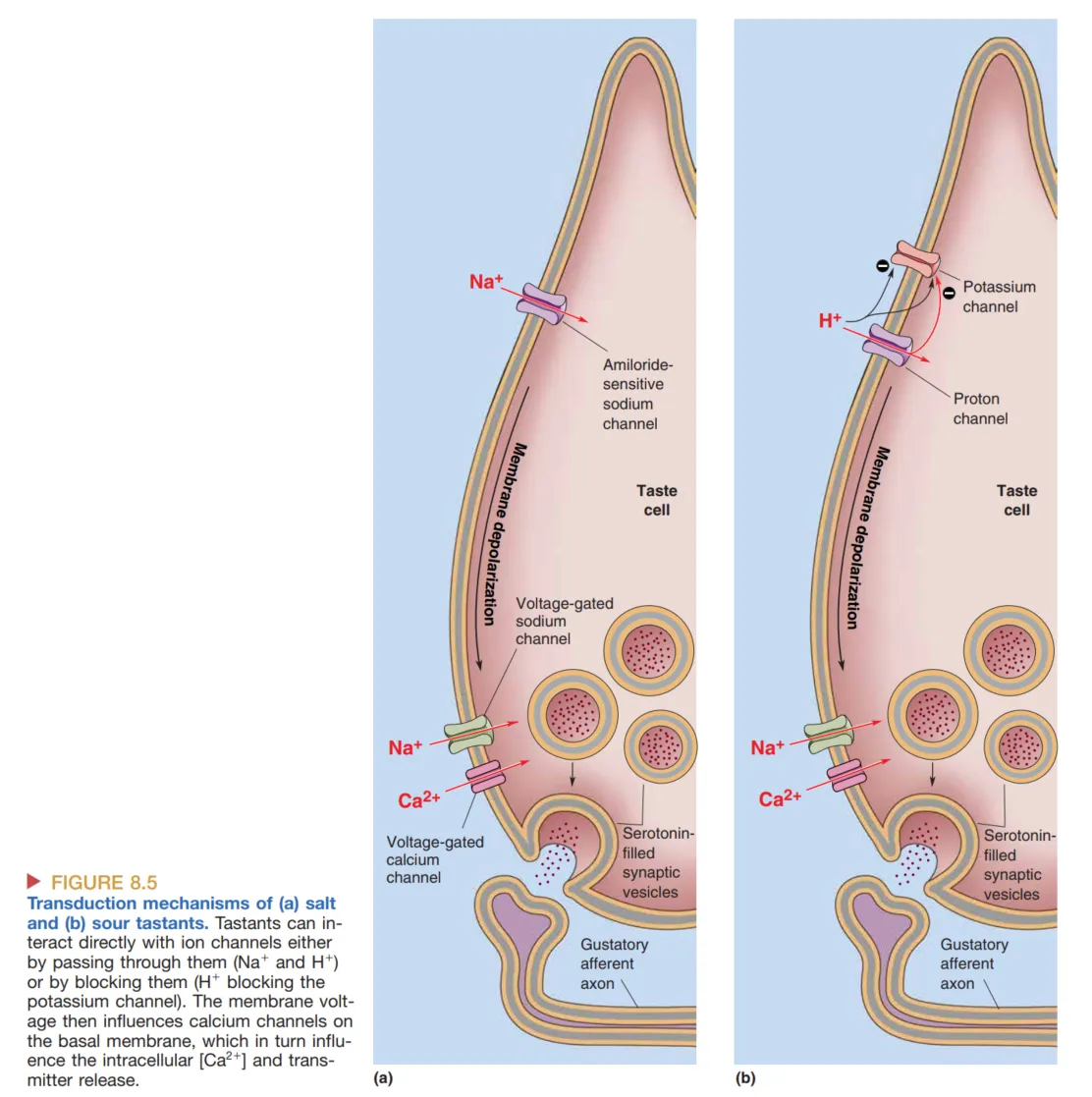
- Saltiness (咸味):主要由 介导。在低浓度下,味觉细胞利用特异性的 Amiloride-sensitive Channels。这种通道对电压不敏感且常开。当外界 浓度升高,离子顺电化学梯度内流,导致膜去极化。在大剂量食盐引起的咸味感知中,阴离子(如醋酸盐)可能会由于空间位阻或电荷效应调节 的效应,其具体机制尚在研究中。
- Sourness (酸味):由高浓度的 (低 pH 值)引起。其机制包括 通道的直接内流以及 对特异性 选透性通道 的结合与阻断。此外,一些 TRP (Transient Receptor Potential) 系列通道也可能参与介导质子带动的去极化。
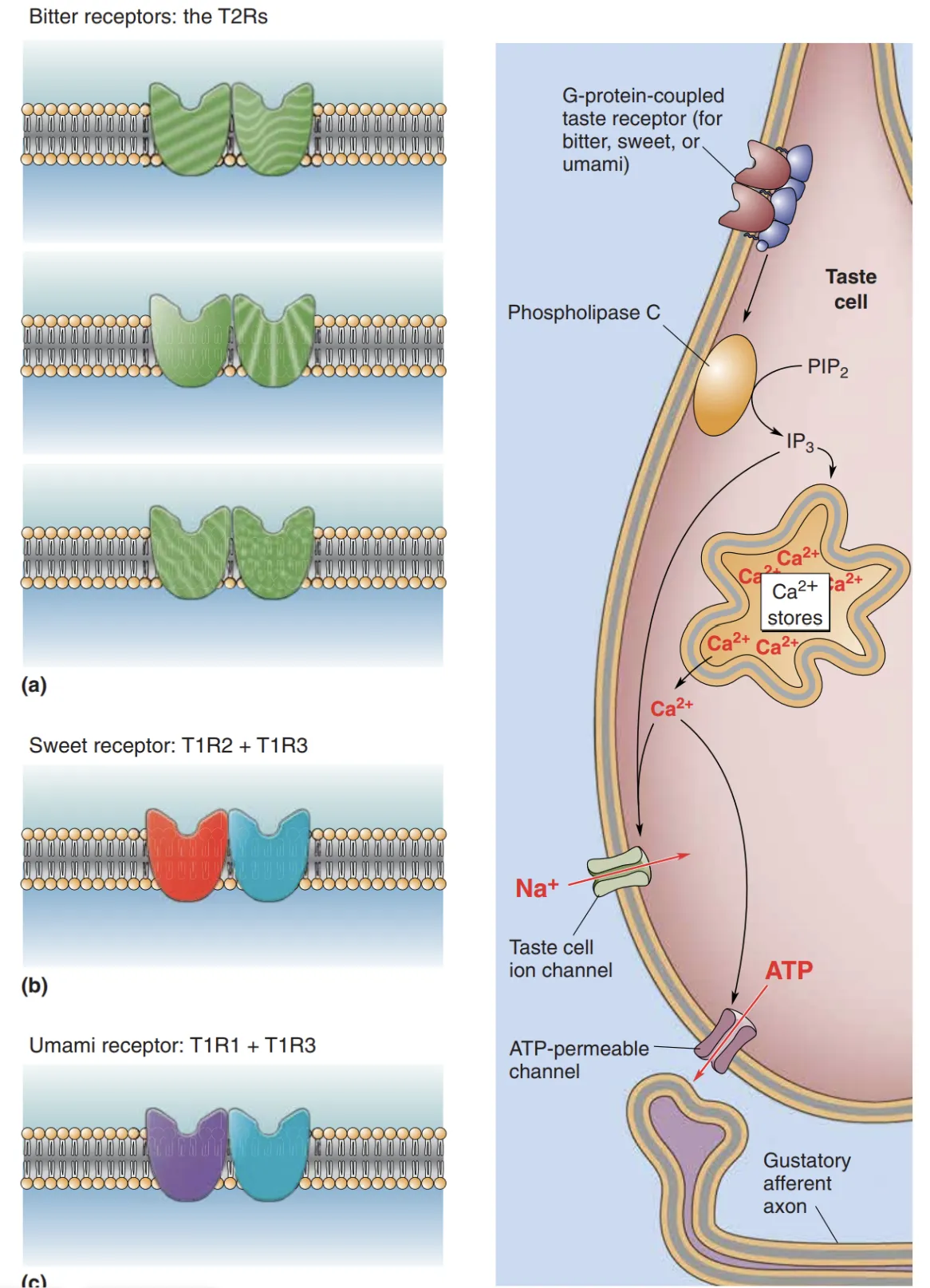
- Bitterness, Sweetness, and Umami (苦、甜、鲜):这三者均使用 G-Protein-Coupled Receptors (GPCRs) 家族。
- Receptors: 苦味通过 T2R 家族(约 25 种亚型)感应;甜味通过 T1R2 + T1R3 异源二聚体(Dimer)识别;鲜味则由 T1R1 + T1R3 异源二聚体探测。
- Transduction Cascade: 递质结合受体后,激活 G 蛋白并触发 Phospholipase C (PLC)。PLC 增加第二信使 ,进而导致胞内钙库释放 。与传统神经元不同,这些细胞不使用突触小泡,而是通过钙激活的钙渗透通道让 ATP 直接从胞浆流出,作为神经递质作用于初级传入纤维上的嘌呤受体。
Central Taste Pathways and Neural Coding
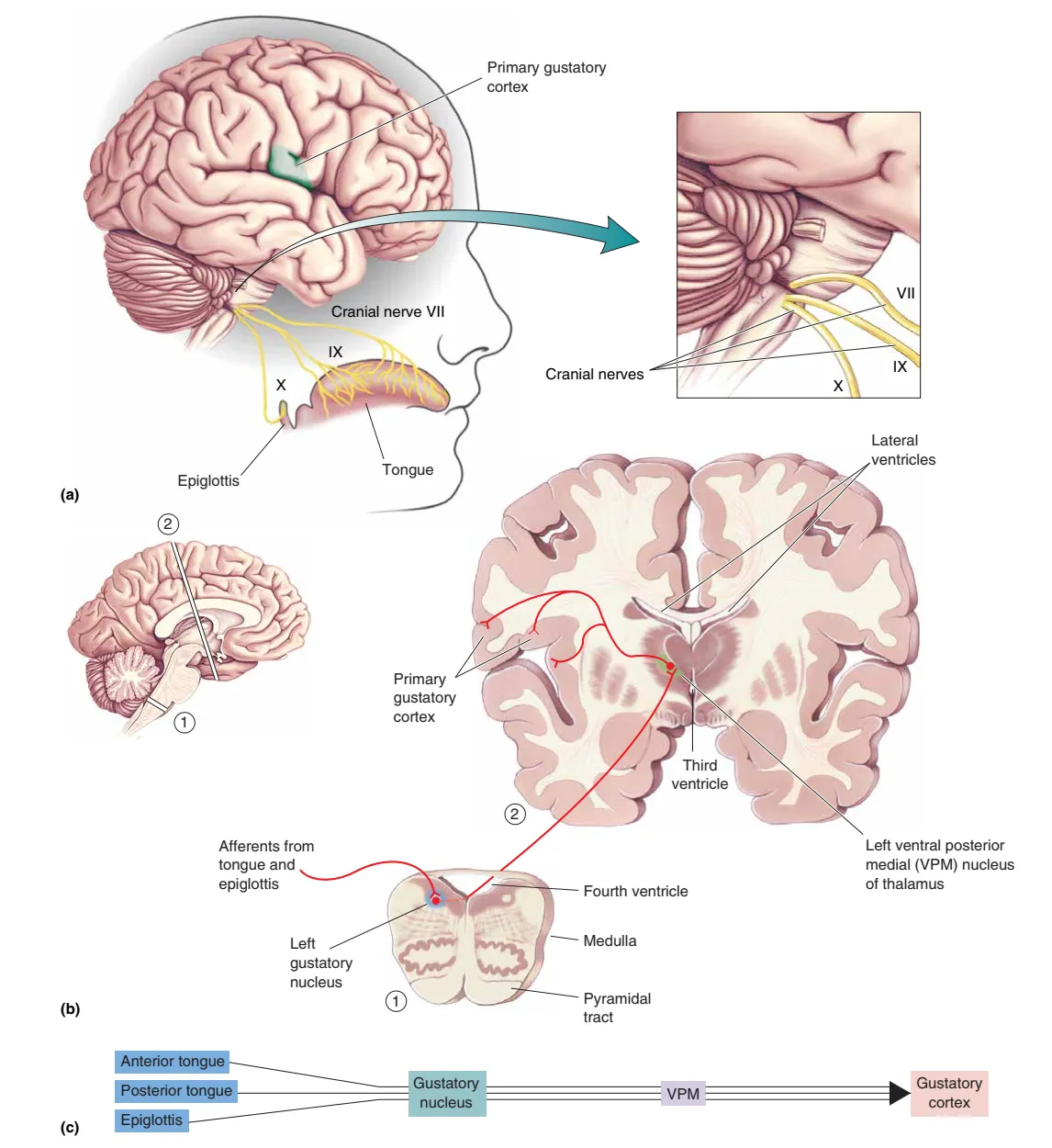
味觉信息通过三对脑神经传递:舌前 2/3 由 VII (Facial, 面神经) 支配;舌后 1/3 由 IX (Glossopharyngeal, 舌咽神经) 支配;口咽及会厌部则由 X (Vagus, 迷走神经) 传递。这些轴突均投射至大脑延髓的 Solitary Nucleus (孤束核) 内的味觉核。随后,神经元继续向上投射至丘脑的 VPM Nucleus (丘脑腹后内侧核),最终到达位于岛叶 (Insula) 的初级味觉皮层。该路径通常是 Ipsilateral (同侧的),相关损伤可能导致 Ageusia (失味症)。
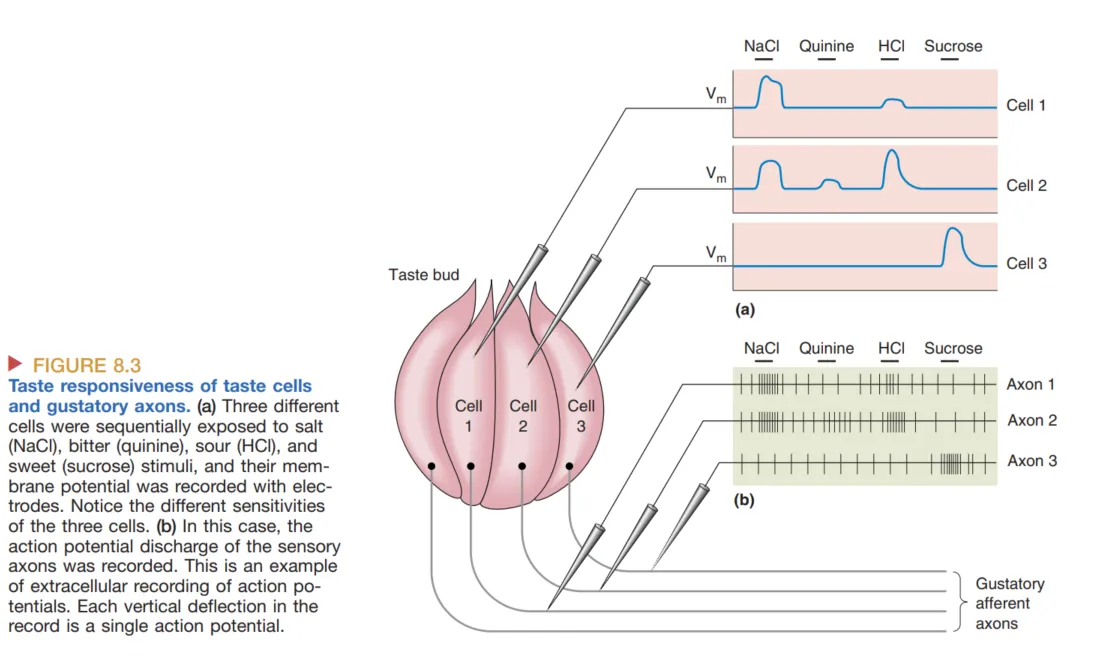
 在神经编码方面,传统的 Labeled Line Hypothesis(标记线假说)认为特定通道专门传递特定味道。然而,现代观点更倾向于 Population Coding (群体编码) 或者标记线与群体编码的融合:大部分味觉神经元是广谱反应(Broadly Tuned)的,大脑通过解读大规模神经元群体的组合活动模式(而非单一细胞的激活)来精确区分食物的精细风味。
在神经编码方面,传统的 Labeled Line Hypothesis(标记线假说)认为特定通道专门传递特定味道。然而,现代观点更倾向于 Population Coding (群体编码) 或者标记线与群体编码的融合:大部分味觉神经元是广谱反应(Broadly Tuned)的,大脑通过解读大规模神经元群体的组合活动模式(而非单一细胞的激活)来精确区分食物的精细风味。
Olfaction (嗅觉)
The Olfactory Apparatus
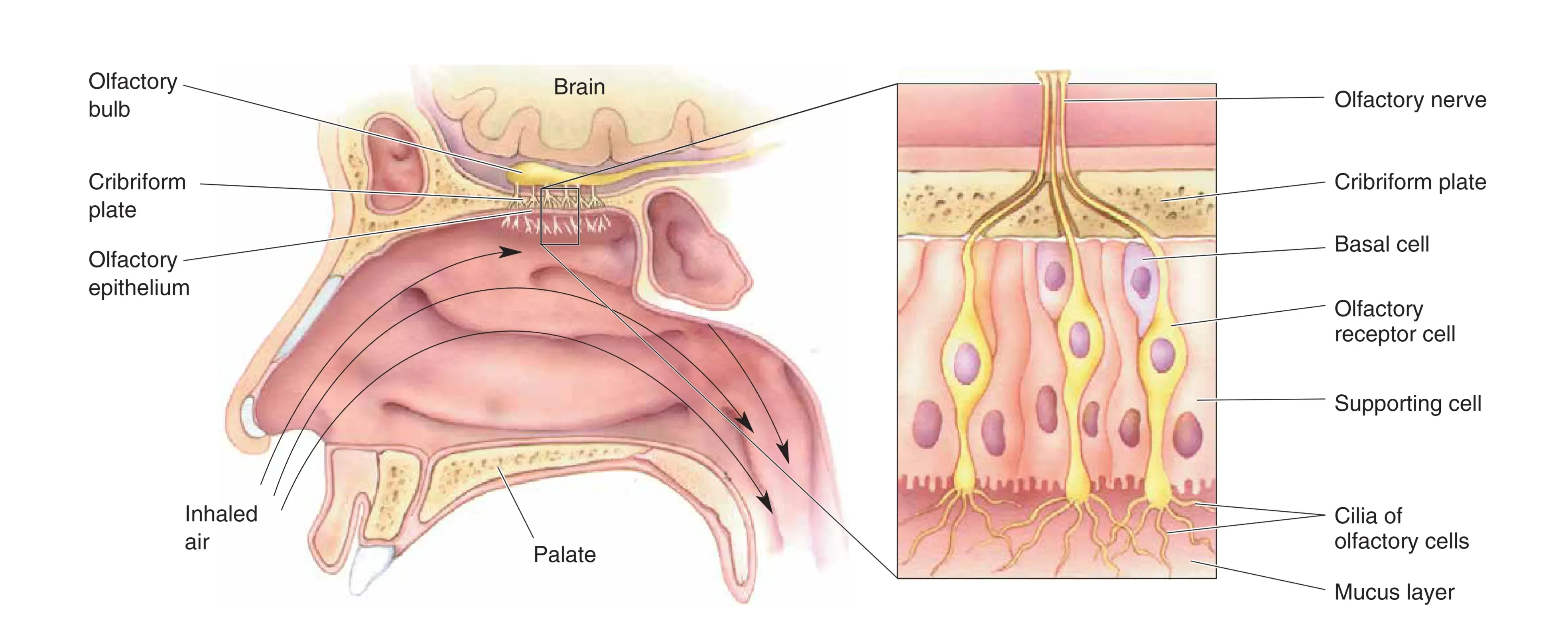
我们并非直接用鼻子闻气味,而是依靠鼻腔顶部的 Olfactory Epithelium (嗅上皮)。它包含三种核心细胞:
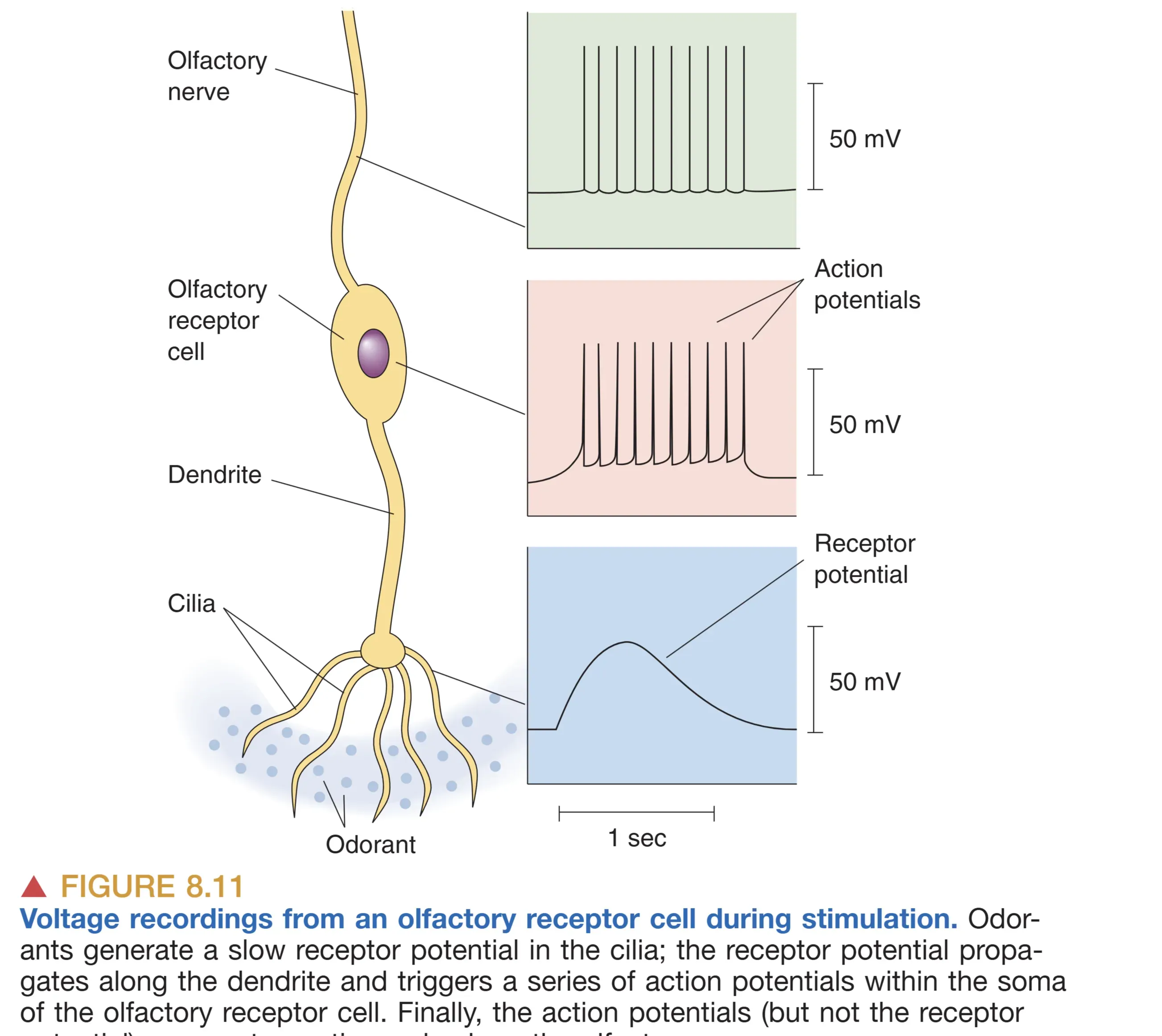
- Olfactory Receptor Cells (嗅觉受体细胞):这是真正的神经元。其纤毛伸入黏液层,轴突穿过 Cribriform Plate (筛板) 进入中枢。其轴突极其纤细且无髓鞘,合称嗅神经(I 对脑神经)。
- Supporting Cells:功能类似胶质细胞,负责产生 Mucus (黏液)(每 10 分钟替换一次,含抗体、酶及嗅质结合蛋白)。
- Basal Cells:干细胞,提供嗅觉神经元持续的再生(周期约为 4-8 周)。
头部外伤若震裂筛板,可能剪断脆弱的嗅觉轴突导致 Anosmia (失嗅症)。此外,除了普通气味,某些生物还通过犁鼻器(Vomeronasal organ)探测 Pheromones (信息素),虽然人类的这一功能高度退化,但气味对情绪和周期的影响依然存在。
Olfactory Transduction Mechanisms
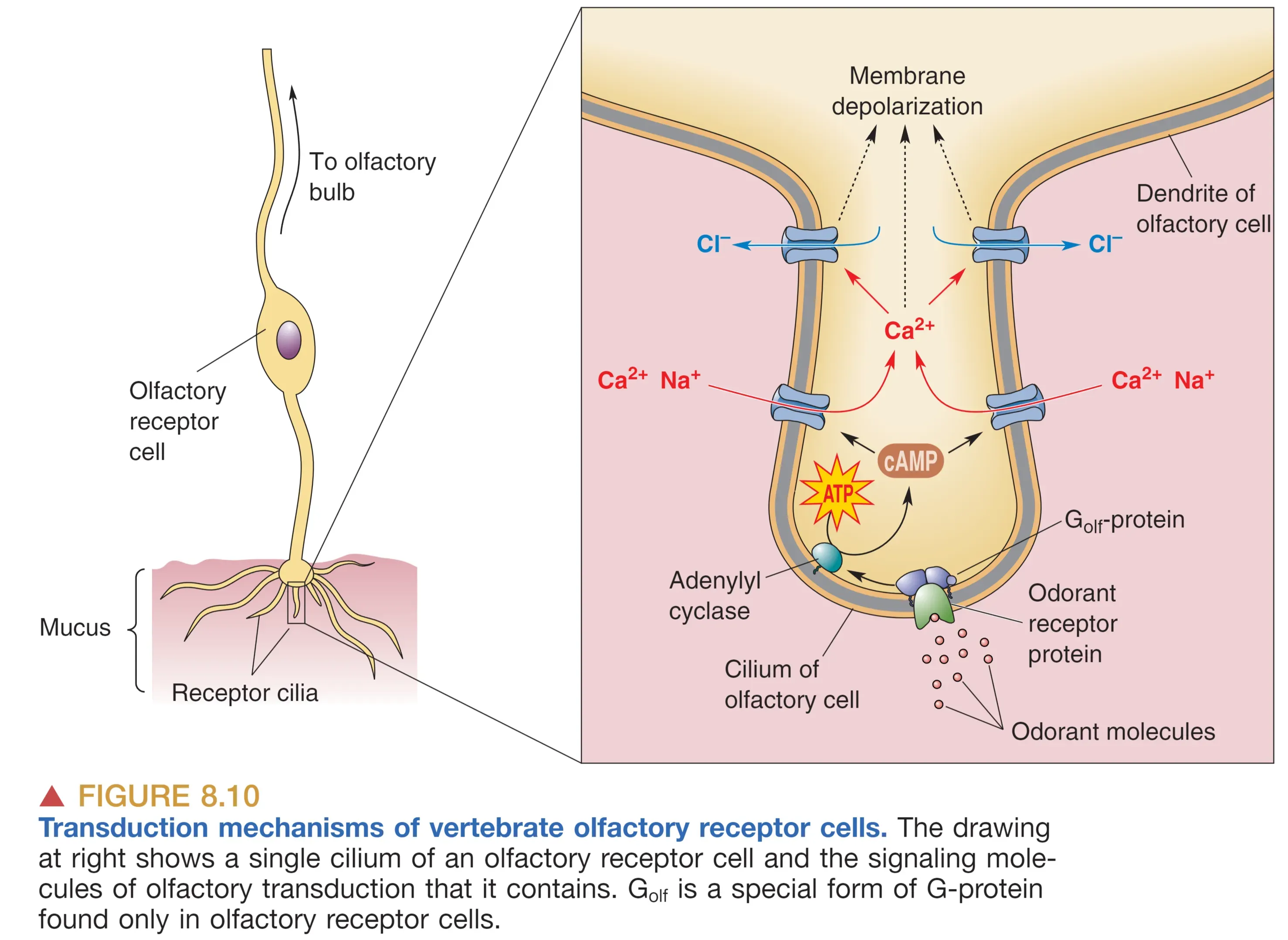
嗅觉转导几乎完全依赖于 cAMP 第二信使通路。
- 气味分子(Odorants)溶解于黏液并结合受体。
- 激活特异性 G 蛋白 ,进而激活腺苷酸环化酶(III型)。
- cAMP 升高直接开启 cAMP-gated Channels,允许 和 内流导致去极化。
- 关键放大与独特性: 调高的胞内 开启了 -activated channels。在嗅觉神经元中,胞内 浓度异常高,因此 流出细胞,产生进一步的去极化(这与神经系统中 流入导致的抑制作用截然相反)。
即便刺激持续,嗅觉也会发生 Adaptation (适应),这涉及信号通路的反馈性下调。信号的终结则通过扩散、黏液层中的酶降解以及胞内反馈机制共同达成。
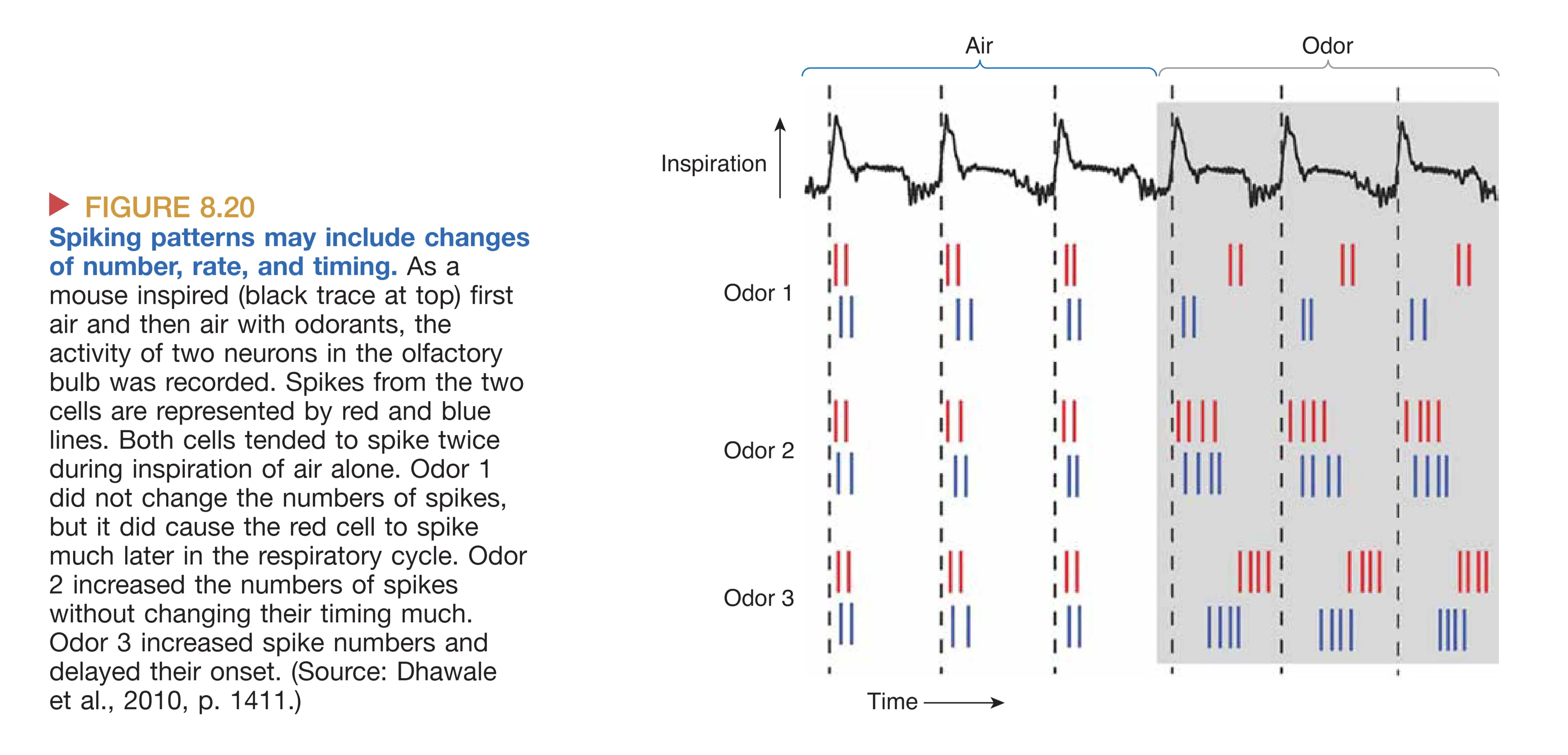
 如果嗅觉受体细胞产生的电位足够大,超过胞体产生动作电位的阈值,则产生的spike可以通过嗅神经传导到中枢神经系统,最终引起嗅觉。
如果嗅觉受体细胞产生的电位足够大,超过胞体产生动作电位的阈值,则产生的spike可以通过嗅神经传导到中枢神经系统,最终引起嗅觉。
Coding and Central Processing
Linda Buck 和 Richard Axel 的研究揭示了哺乳动物拥有庞大的嗅觉受体基因家族(人类约有 350 种表达基因)。遵循 “One Cell, One Receptor” 原则,每个受体细胞通常仅表达一种受体蛋白。
- Olfactory Bulb and Glomeruli: 嗅神经纤维会聚到嗅球中的 Glomeruli (嗅小球)。映射极其精准:表达相同受体基因的数万个神经元会聚到极少数(约 2000 个)特定的嗅小球上。
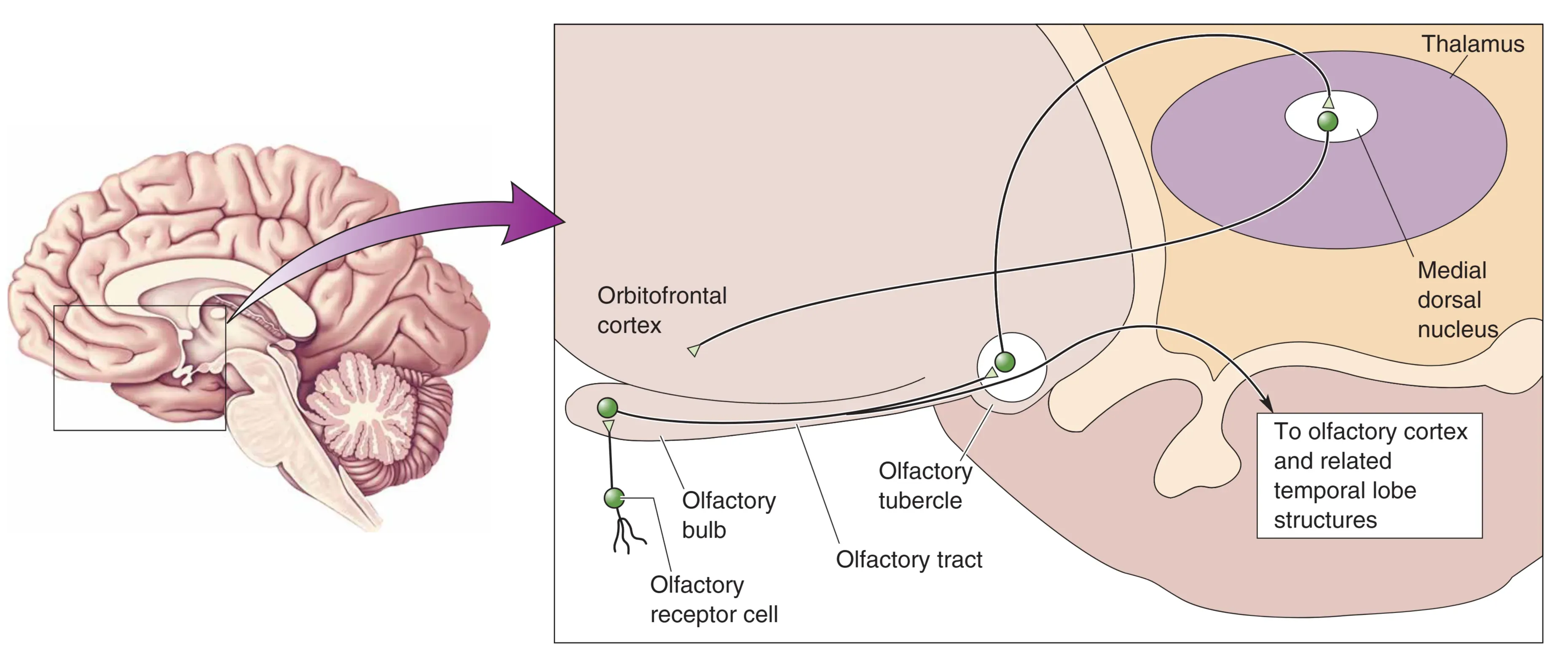
- Central Pathways: 嗅觉是唯一不需要先经过丘脑换元、直接投射到皮层的感觉。
- Pathway A: 嗅球 -> 嗅结节 -> 丘脑 MD Nucleus -> Orbitofrontal Cortex (介导意识感知)。
- Pathway B: 嗅球 -> 直接投射到 Piriform Cortex(梨状皮层)和边缘系统(杏仁核等),这解释了为什么气味能强烈唤起情感与记忆。
Spatial and Temporal Representations
嗅觉解码依赖三种策略:
- Population Coding: 即使单细胞是广谱的,成千上万个细胞的组合提供了精确的方向。
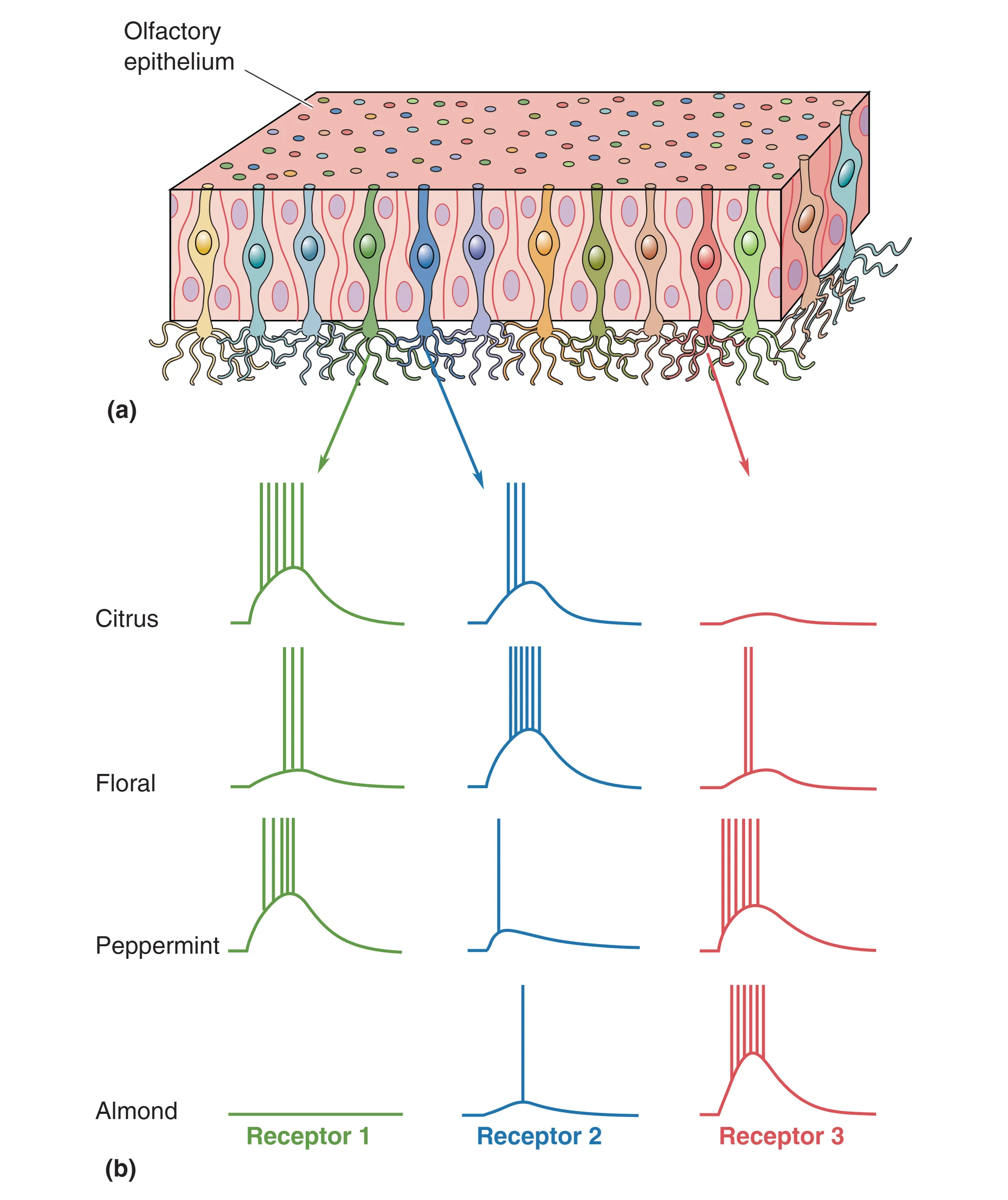
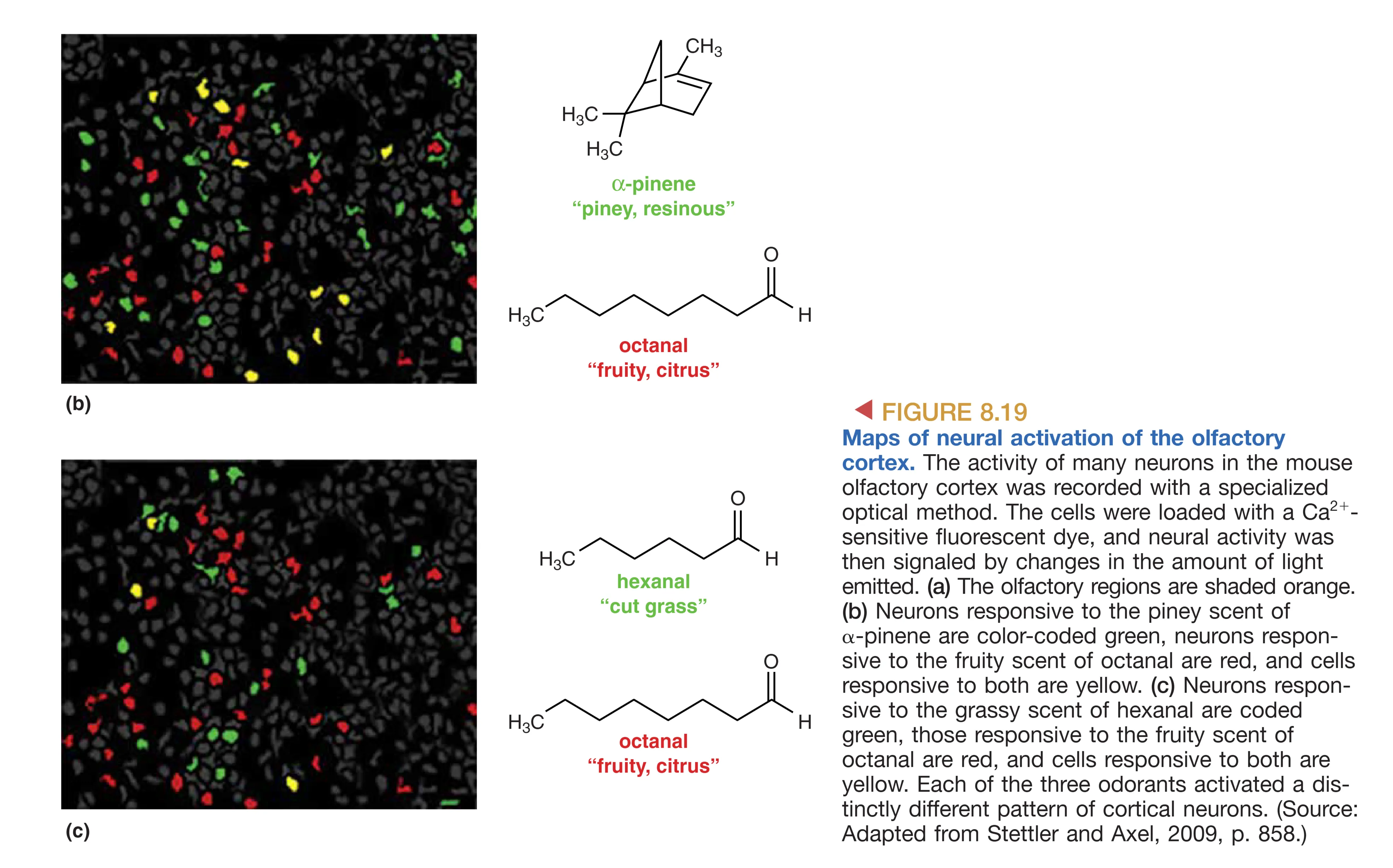
- Sensory Maps (Spatial Maps):嗅球中小球的激活模式形成了一种空间化的“气味地图”。
- Temporal Coding: 动作电位的时间节律、放电频率以及细胞间的同步性携带了气味质量的关键信息。值得一提的是,语言处理偏向左脑,而嗅觉处理往往在右脑表现出更强的偏向性。